- 東京都健康長寿医療センター(研究所)
- English
- Research Team
- Research Team for Aging Science
- Muscle Biology
Muscle Biology
Members
| Theme Leader | Yusuke Ono, Ph.D. |
| Researchers | Kodai Nakamura, Ph.D., Takashi Sasaki, Ph.D. |
| Technician | Kanan Iwata |
Keywords
Sarcopenia, Muscular dystrophy, Muscle plasticity, Muscle regeneration, Satellite cells, Exercise, Muscle memory, FSHD, DUX4, Dll4, Notch signaling
Major Research Titles
- Understanding of molecular mechanisms underlying muscle resilience, plasticity and diversity
- Development of molecular therapies for muscle wasting diseases
Profile
Skeletal muscle is the largest tissue in the adult human body accounting for approximately 40% of the body weight, responsible for movement and locomotion by generating contraction forces. Skeletal muscle is a highly plastic tissue that functionally adapts its structure and metabolism in response to chemical and mechanical stimuli. While high intensity resistance training induces muscle hypertrophy, mechanical unloading such as immobilization and disuse results in muscle atrophy. Satellite cells are the muscle stem cells that are located between the basal lamina and the plasmalemma of myofibers, providing new myonuclei for postnatal muscle growth and hypertrophy, and regeneration in adult muscle. We are interested in understanding the cellular and molecular mechanisms that control muscle resilience, plasticity and diversity. Our goal is to develop new strategies to treat muscle wasting diseases including muscular dystrophy and age-related sarcopenia.
Recent Publications
Nakamura et al., J Clin Invest. 2025 Jul 1;135(17):e181881.
Iron supplementation improves muscle function in a mouse model of muscular dystrophy
Facioscapulohumeral muscular dystrophy (FSHD), a rare genetic muscle disease. FSHD is characterized by progressive muscle weakness that typically begins in the face and shoulders and gradually spreads to the upper arms and lower limbs. The disease
is caused by aberrant expression of the toxic transcription factor DUX4 in skeletal muscle, which induces oxidative stress, inflammation, and muscle degeneration. Although DUX4 is widely recognized as a central therapeutic target, the molecular processes
that translate DUX4 expression into muscle damage remain incompletely understood.
In the present study, we focused on iron metabolism, a critical regulator of oxidative stress. Using a genetically engineered mouse model that conditionally expresses DUX4 in skeletal muscle, we found that DUX4 disrupted intracellular iron homeostasis,
leading to iron accumulation in muscle tissue. This abnormal iron accumulation resulted in oxidative damage and activation of ferroptosis pathway, which is known as an iron-mediated cell death driven by excessive lipid peroxidation. Importantly, iron
supplementation--administered through diet as well as via ferric carboxymaltose (FCM), an FDA-approved intravenous iron formulation--significantly reduced pathological iron accumulation in muscle tissue. Treated mice showed marked improvements in muscle
structure and function, including increased grip strength, enhanced muscle force generation, and improved treadmill running performance. Transcriptome analyses further revealed that iron supplementation suppressed the abnormal activation of inflammatory
and lysosomal pathways induced by DUX4, providing mechanistic insight into how restoring iron balance protects muscle tissue from degeneration. Furthermore, in vitro compound library screening uncovered a drug to attenuate DUX4 toxicity, which was ferrostatin-1
(Fer-1), a potent inhibitor of ferroptosis. Treatment with Fer-1 in DUX4-Tg mice successfully improved grip strength and running performance, suggesting that the ferroptosis pathway could be a therapeutic target for FSHD. Although clinical studies will
be required to confirm safety and efficacy in patients, this work opens a new avenue for FSHD treatment strategies and underscores the potential of repurposing existing iron formulations for neuromuscular diseases.
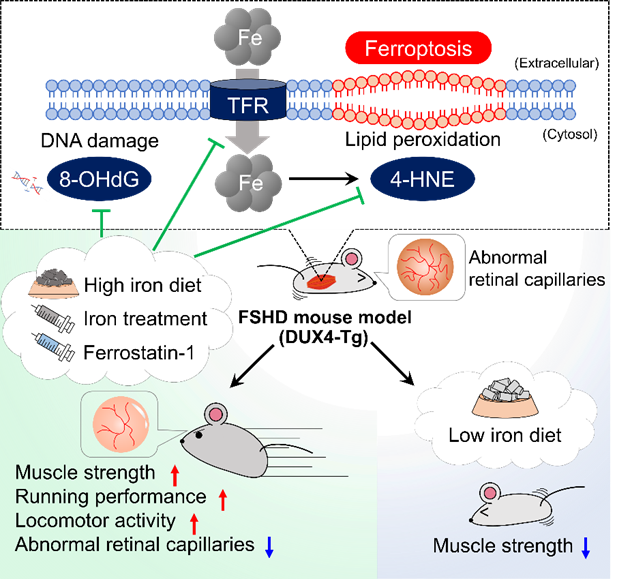 Activation of ferroptosis pathways in FSHD pathogenesis
Activation of ferroptosis pathways in FSHD pathogenesisFujimaki et al., Nature Metab. 2022 Feb;4(2):180-189
Discovery of the vascular Dll4-muscular Notch2 axis in muscle atrophy models
Skeletal muscle is a highly plastic tissue that functionally adapts its metabolism and readily reduces or gains its mass in response to environmental stimuli such as mechanical unloading or overloading. Skeletal muscle also regenerates after injury, and this regenerative capacity relies on the resident muscle tissue stem cells, namely satellite cells. Satellite cells provide myonuclei not only for regeneration but also for postnatal muscle growth and hypertrophy. Notch signaling is evolutionally conserved and regulates the stem cell functions through cell-cell contact during tissue development and regeneration. We have recently reported that satellite cells express Notch1 and Notch2, which are essential for muscle regeneration by coordinately maintaining the quiescent and undifferentiated state of satellite cells in adult mice. Further analysis revealed that Notch2 is expressed not only in satellite cells but also in terminally differentiated multinucleated myofibers, acting as a key mediator in mechanical unload (disuse)- or metabolic overload (hyperglycemia)-induced muscle atrophy. Mechanistically, in both atrophic conditions, the microvascular endothelium upregulates and releases a Notch ligand Delta-like ligand-4 (Dll4) as a soluble form, which then activates muscular Notch2 without affecting satellite cell function. Inhibition of the Dll4-Notch2 axis substantively prevents muscle atrophy and promotes mechanical overloading-induced muscle hypertrophy in mice. Our results illuminate a tissue-specific function of the microvascular endothelium in controlling tissue plasticity and highlight the vascular Dll4-muscular Notch2 axis as a central upstream mechanism that integrates catabolic signals from mechanical and metabolic stimulation.
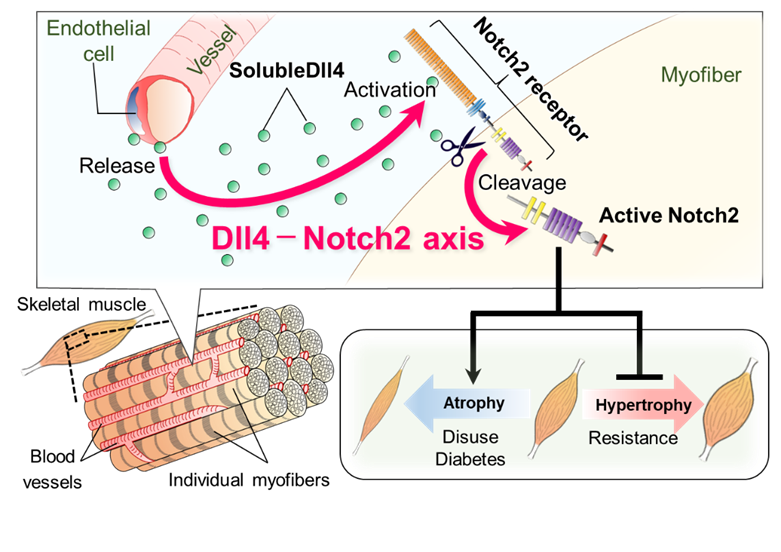 Identification of the vascular Dll4-muscular Notch2 axis
Identification of the vascular Dll4-muscular Notch2 axisYoshioka et al., Science Adv. 2021 Jun 9. 7: eabd7924.
Muscles retain positional memory from fetal life
There are various types of the intractable muscle disease muscular dystrophy and each type has a different symptom location. Similarly, age-related muscle fragility (sarcopenia) does not occur evenly throughout the body. The physical location of
the symptoms of these diseases cannot be explained by differences in muscle fiber types or physical activity patterns alone, and requires a new perspective to elucidate their respective pathogeneses.
The developmental origin of cells that form muscles differ in the fetal stage. For example, most of the craniofacial muscles originate from the cranial mesoderm, while the limb muscles originate from the body segments. Development of limb and craniofacial
muscles in the fetal period involves specific molecular mechanisms that depends on their origin. However, differences in the properties of mature skeletal muscle depending on body position after birth have not been fully discussed. Thus, a research collaboration
worked to visualize the body's positional information by studying the epigenomic state and gene expression patterns of skeletal muscle and the muscle stem cells responsible for regeneration.
Using skeletal muscle and associated muscle stem cells isolated from the heads and hind limbs of adult mice, we investigated positional specificity at the epigenomic level using DNA methylome analysis. They found characteristic differences in the
DNA methylation status at the homeobox (Hox) loci. Among four regions, A to D, the Hox-A locus in particular had an overall DNA hypermethylation state in hindlimb skeletal muscle and muscle stem cells compared to the head. Additionally, both skeletal
muscle and muscle stem cells in the hind limbs showed high expression of the Hox-A gene. Many of these Hox-A genes reflected expression patterns in the fetal period. These findings suggest that skeletal muscle and muscle stem cells remember positional
information during fetal life, and that epigenomic regulation by DNA methylation may be involved in positional memory.
We then focused on the Hoxa10 gene, which was highly expressed only in the limb muscles. When hindlimb-derived muscle stem cells expressing Hoxa10 were isolated and transplanted into craniofacial muscles that do not express Hoxa10, Hoxa10 gene
expression became detectable in the craniofacial muscles. In other words, hindlimb-derived muscle stem cells were able to innervate the craniofacial muscle with strong retention of positional memory even after ectopic transplantation.
We then generated mice lacking the Hoxa10 gene in muscle stem cells to analyze its function. A Hoxa10 deficiency severely impaired the regeneration of hindlimb muscles but had no effect on craniofacial muscle regeneration. A detailed investigation
of the mechanism behind the hindlimb muscle regeneration disorder revealed that it is caused by genomic instability due to abnormal chromosome distribution during muscle stem cell division. Furthermore, analysis of human head and leg muscle stem cells
also showed that only leg muscle cells expressed the HOX-A gene and that its inhibition resulted in abnormal cell division, confirming that muscle positional memory is retained in humans and mice.
This research suggests that the positional memory of muscle stem cells based on the position-specific distribution of Hox gene expression may determine the position-specific properties of skeletal muscle, rather than merely persisting from fetal
life.
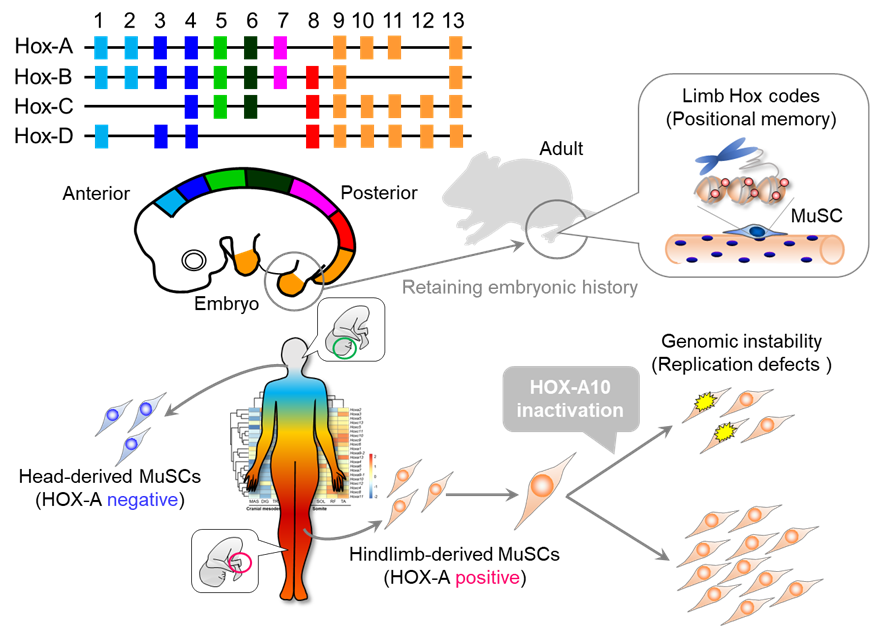 Positional memory in adult skeletal muscle and satellite cells
Positional memory in adult skeletal muscle and satellite cellsTsuchiya et al. Stem Cell Rep. 2020 Oct 13;15(4):926-940.
Damaged muscles don't just die, they regenerate themselves
Skeletal muscle is composed of bundles of contracting myofibers and each myofiber is surrounded by satellite cells that can produce new myofibers. Satellite cells also play essential roles in muscle growth during developmental stages and muscle
hypertrophy during strength training. In mature skeletal muscle, satellite cells are usually present in a dormant state. Upon stimulation after muscle injury, satellite cells are rapidly activated and proliferate extensively. During the subsequent myogenesis,
they differentiate and regenerate muscle fibers by fusing with existing muscle fibers or with together. Of these three steps (satellite cell activation, proliferation, and muscle differentiation), little is known about how the first step, activation,
is induced.
Since satellite cells are activated when muscle fibers are damaged, we hypothesized that muscle damage itself could trigger activation. However, this is difficult to prove in animal models of muscle injury so they constructed a cell culture model
in which single muscle fibers, isolated from mouse muscle tissue, were physically damaged and destroyed. Using this injury model, they found that components leaking from the injured muscle fibers activated satellite cells, and the activated cells entered
the G1 phase of cell division. Further, the activated cells returned to a dormant state when the damaged components were removed, thereby suggesting that the damaged components act as the activation switch.
We named the leaking components "Damaged myofiber-derived factors" (DMDFs), after the broken muscle fibers, and identified them using mass spectrometry. Most of the identified proteins were metabolic enzymes, including glycolytic enzymes such as
GAPDH, and muscle deviation enzymes that are used as biomarkers for muscle disorders and diseases. GAPDH is known as a "moonlighting protein" that has other roles in addition to its original function in glycolysis, such as cell death control and immune
response mediation. We therefore analyzed the effects of DMDFs, including GAPDH, on satellite cell activation and confirmed that exposure resulted in their entry into the G1 phase. Furthermore, we injected GAPDH into mouse skeletal muscle and observed
accelerated satellite cell proliferation after subsequent drug-induced muscle damage. These results suggest that DMDFs have the ability to activate dormant satellite cells and induce rapid muscle regeneration after injury. The mechanism by which broken
muscle activates satellite cells is a highly effective and efficient tissue regeneration mechanism.
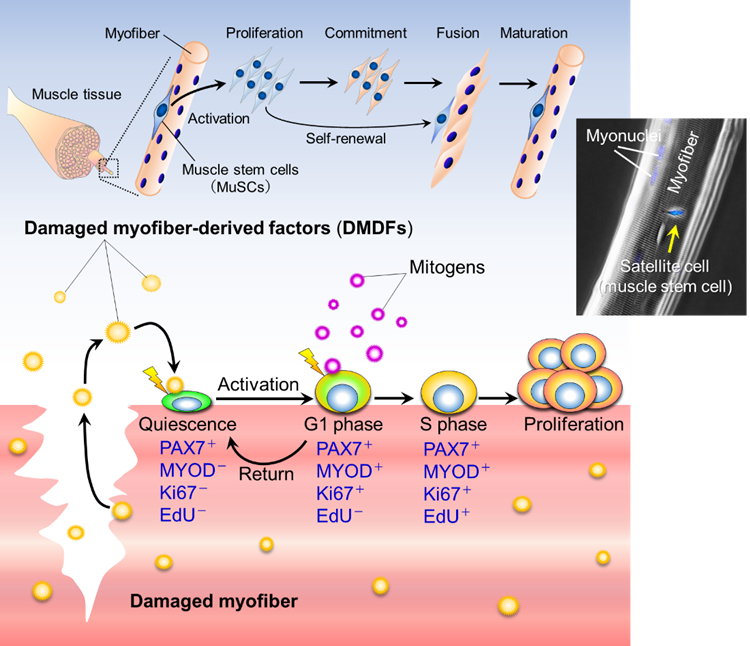 DMDFs activate muscle satellite cells
DMDFs activate muscle satellite cellsSeko and Fujita et al. Stem Cell Rep. 2020 Sep 8;15:577-586.
Skeletal muscle regeneration mechanisms vary by gender
An epidemiological study of pre and postmenopausal women in their 50s indicated an association between decreased blood estrogen levels and muscle weakness. We previously showed that estrogen is important for skeletal muscle development and regeneration
using an ovariectomized estrogen deficiency mouse model (Kitajima and Ono, J Endocrinol 2016). We also examined the effectiveness of nutritional interventions in estrogen-deficient conditions (Kitajima et al., Nutrients 2017). However, whether estrogen
acts directly on the estrogen receptors (ERs) of myofibers and muscle stem cells to regulate muscle growth and regeneration, or whether it acts indirectly through other tissues and organs was unclear. In this study, we generated mice with either myofiber-specific
or muscle stem cell-specific ERβ gene deletion and analyzed the function of ERβ in skeletal muscle.
To clarify the role of ERβ in the growth of skeletal muscle, we generated mice (mKO) in which the action of the ERβ gene could be turned off in myofibers with the administration of the drug doxycycline. ERβ deficiency was induced at 6 weeks of
age, and muscle fiber area and strength of the tibialis anterior muscle was measured at 10-12 weeks. Compared to control mice, both indices were reduced in female mKO mice but not in male mice. Since there was no change in the expression of muscle atrophy-related
genes, this reduced growth of female mice was not thought to be due to an increase in muscle atrophy.
To determine the function of ERβ in muscle stem cells, we generated scKO mice in which the ERβ gene could be deleted in muscle stem cells with the administration of the drug tamoxifen. They then evaluated muscle regenerative capacity by locally
inducing muscle damage. While muscle regeneration was efficient in control mice, the regenerated muscle tissue of female scKO mice showed thin regenerated muscle fibers, fibrosis caused by collagen deposition, and significantly reduced muscle regenerative
capacity. Muscle regeneration in male scKO mice, however, was not impaired. Because impaired muscle regeneration in females was not exacerbated by ovariectomies that made them estrogen deficient, we thus thought that estrogen regulates muscle regeneration
via ERβ expressed by muscle stem cells. To further investigate the cause of reduced muscle regenerative capacity, we isolated and cultured muscle stem cells for evaluation. ERβ in cells from scKO mice was evaluated in several experiments using siRNAs
and inhibitors. ERβ was found to contribute to the promotion of muscle stem cell proliferation and the inhibition of cell death. RNA-seq of scKO muscle stem cells showed that the expression of "niche"-related genes, which are required for the maintenance
of stem cell properties, was reduced in scKO muscle stem cells. Therefore, we speculate that the inactivation of ERβ may have affected the proliferation and survival of muscle stem cells by inhibiting the formation of stem cell niches.
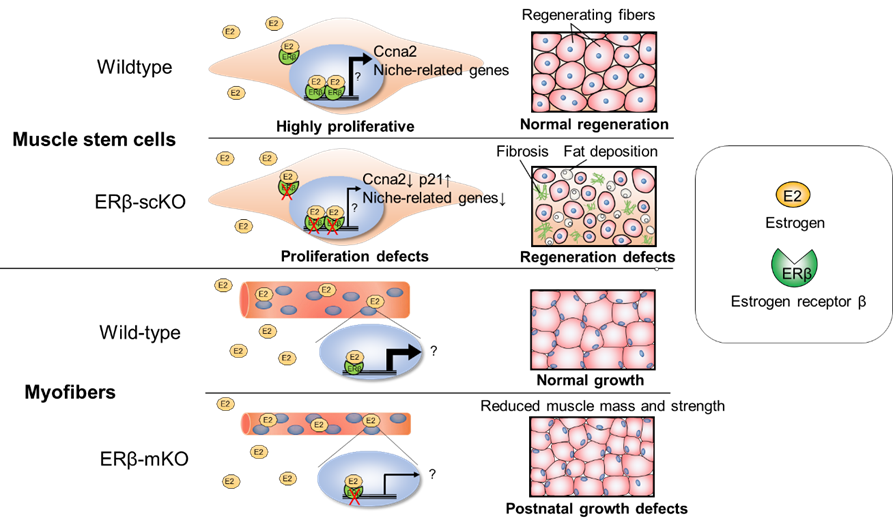 ERβ controls muscle growth and regeneration
ERβ controls muscle growth and regenerationReferences
Major Publications
Original Articles
- Nakamura K, Ortuste-Quiroga HP, Horii N, Fujimaki S, Moroishi T, Nakayama KI, Hino S, Saito Y, Nishino I, Ono Y. Iron supplementation alleviates pathologies in a mouse model of facioscapulohumeral muscular dystrophy. J Clin Invest. 2025 Jul 1;135(17):e181881.
- Kitajima Y, Yoshioka K, Mikumo Y, Ohki S, Maehara K, Ohkawa Y, Ono Y. Loss of Tob1 promotes muscle regeneration through muscle stem cell expansion. J Cell Sci. 2024 Aug 1;137(15):jcs261886.
- Fujimaki S, Ono Y. Murine models of tenotomy-induced mechanical overloading and tail-suspension-induced mechanical unloading. Methods Mol Biol. 2023 Mar 30; 2640:207-215.
- Fujimaki S, Matsumoto T, Muramatsu M, Nagahisa H, Horii N, Seko D, Masuda S, Wang X, Asakura Y, Takahashi Y, Miyamoto Y, Usuki S, Yasunaga KI, Kamei Y, Nishinakamura R, Minami T, Fukuda T, Asakura A, Ono Y. The endothelial Dll4−muscular Notch2 axis regulates skeletal muscle mass. Nature Metab. 2022 Feb;4(2):180-189.
- Tsuchiya Y, Ono Y. An in vitro mechanical damage model of isolated myofibers in a floating culture condition. Bio Protoc. 2022 Jan 5;12(1):e4280.
- Yoshioka K, Nagahisa H, Miura F, Araki H, Kamei Y, Kitajima Y, Seko D, Nogami J, Tsuchiya Y, Okazaki N, Yonekura A, Ohba S, Sumita Y, Chiba K, Ito K, Asahina I, Ogawa Y, Ito T, Ohkawa Y, Ono Y. Hoxa10 mediates positional memory to govern stem cell function in adult skeletal muscle. Science Adv. 2021 Jun 9. 7: eabd7924.
- Yoshioka K, Kitajima Y, Seko D, Tsuchiya Y, Ono Y. The body-region-specificity in murine models of muscle regeneration and atrophy. Acta Physiol (Oxf). 2021 Jan;231(1):e13553.
- Tsuchiya Y, Kitajima Y, Masumoto H, Ono Y. Damaged myofiber-derived metabolic enzymes act as activators of muscle satellite cells. Stem Cell Rep. 2020 Oct 13;15(4):926-940. Highlighted Featured Article/Most Read in Stem Cell Reports (The Best of Stem Cell Reports 2020−2021).
- Kitajima Y, Suzuki N, Yoshioka K, Izumi R, Tateyama M, Tashiro Y, Takahashi R, Aoki M, Ono Y. Inducible Rpt3, a proteasome component, knockout in adult skeletal muscle results in muscle atrophy. Front Cell Dev Biol. 2020 Sep 2;8:859.
- Seko D, Fujita R, Kitajima Y, Nakamura K, Imai Y, Ono Y. Estrogen receptor β controls muscle growth and regeneration in young female mice. Stem Cell Rep. 2020 Sep 8;15:577-586. Highlighted. Featured Article in Stem Cell Reports.
- Yoshioka K, Kitajima Y, Okazaki N, Chiba K, Yonekura A, Ono Y. A Modified pre-plating method for high-yield and high-purity muscle stem cell isolation from human/mouse skeletal muscle tissues. Front Cell Dev Biol. 2020 Aug 13;8:793.
- Yoshioka K, Fujita R, Seko D, Suematsu T, Miura S, Ono Y. Distinct roles of Zmynd17 and PGC1α in mitochondrial quality control and biogenesis in skeletal muscle. Front Cell Dev Biol. 2019 Dec 10;7:330.
- Kitajima Y, Suzuki N, Nunomiya A, Osana S, Yoshioka K, Tashiro Y, Takahashi R, Ono Y, Aoki M, Nagatomi R. The Ubiquitin-proteasome system is indispensable for the maintenance of muscle stem cells. Stem Cell Rep. 2018 Dec 11;11(6):1523-1538. Highlighted.
- Fujita R, Yoshioka K, Seko D, Suematsu T, Mitsuhashi S, Senoo N, Miura S, Nishino I, Ono Y. Zmynd17 controls muscle mitochondrial quality and whole-body metabolism. FASEB J. 2018 Sep;32(9):5012-5025.
- Kitajima Y and Ono Y. Visualization of PAX7 protein dynamics in muscle satellite cells in a YFP-knock-in-mouse line. Skelet Muscle. 2018 Aug 24;8(1):26.
- Fujimaki S, Seko D, Kitajima Y, Yoshioka K, Tsuchiya Y, Masuda S, Ono Y. Notch1 and Notch2 coordinately regulate muscle stem cell function in the quiescent and activated states. Stem Cells. 2018 Feb;36(2):278-285.
- Kitajima Y, Ogawa S, Egusa S, Ono Y, Soymilk improves muscle weakness in young ovariectomized female mice. Nutrients. 2017 Aug 4;9(8). pii: E834.
- Kitajima Y, Ogawa S and Ono Y. Visualizing the functional heterogeneity in muscle stem cells. Methods Mol Biol. 2016 1516:183-93.
- Kitajima Y, Ono Y. Estrogens maintain skeletal muscle and satellite cell functions. J Endocrinol. 2016 Jun. 229, 267-275.
- Seko D, Ogawa S, Li TS, Taimura A, Ono Y, μ-Crystallin controls muscle function through thyroid hormone action. FASEB J. 2016 May;30(5):1733-40.
- Masuda S, Hisamatsu T, Seko S, Urata Y, Goto S, Li TS, Ono Y. Time- and dose-dependent effects of total-body radiation on muscle stem cells. Physiol Rep. 3 (4), 2015, e12377.
- Ono Y, Urata Y, Goto S, Nakagawa S, Humbert PO, Li TS, Zammit PS. Muscle stem cell fate is controlled by the cell polarity protein Scrib. Cell Rep. 2015 Feb 24;10(7):1135-48.
Review Articles
- Okino R, Goda, Y, Ono Y. The Hox-based positional memory in muscle stem cells. J Biochem. 2024 Aug 28:mvae059. Review Article.
- Ortuste Quiroga HP, Fujimaki S, Ono Y. Pax7 reporter mouse models: a pocket guide for satellite cell research. Eur J Trans Myol. 2023 Dec 18. Review Article.
- Fujimaki S and Ono Y. Notch signaling in the regulation of skeletal muscle stem cells. J Phys Fitness Sports Med, 7 (4): 213-219 (2018) Review Article.
- Fujita R and Ono Y. eIF2α, a potential target for stem cell-based therapies. Stem Cell Investigation, 2016 2016 Jul 22;3:30. Editorial.
- Fujita R, Seko S, Ono Y. Scribble dictates orderly stem cell fate. Oncotarget, 2015, Vol. 6, No.22.18738-9. Editorial.
- Ono Y. Satellite cell heterogeneity and hierarchy in skeletal muscle. J Phys Fitness Sports Med, 3(2): 229-234, 2014. Review Article.